記憶・学習研究グループ
 2.感覚野(皮質)−海馬の相互作用と記憶の理論モデル
2.感覚野(皮質)−海馬の相互作用と記憶の理論モデル
皮質の記憶システムは2種の入力を受けます。1つは感覚野からの事象情報(local、bottom up)であり、もうひとつは海馬からの文脈情報(global、top down)です。この2つの入力は皮質において動的に相互作用し、その相互作用した情報は長期記憶として皮質に蓄えられます。我々はいかにそれらの入力が聴覚皮質上で相互に作用するかを調査するために、海馬の電気刺激と純音提示をペアリングしました。この実験において、聴覚野の活動を光学計測しました。この結果、聴覚皮質に音刺激と海馬刺激間のタイミングに依存したゲーテング機構が存在し、タイミングに依存して聴覚野の神経活動が修飾されます。その修飾は海馬刺激の強弱に依存して抑制性(弱)から興奮性(強)に変化しました。
海馬-皮質間の記憶モデル
生理学実験における上記証拠に基づき、理論的な海馬-皮質記憶モデルを我々は提案しました(Pan and Tsukada, 2006)。塚田らは、生理実験に基づいた海馬‐皮質系のモデルを提案し文脈に基づく記憶書き込み、および読み出しのモデルを実現しています。このモデルはつぎの2段階の機能からなります。
第1段階:塚田らの提案した時空間学習則が有効に働き、海馬神経回路網に時空間の出来事の文脈をつくる。
第2段階:ヘブの学習則により連合野に出来事の類似性の構造(距離空間)をもつ強固なアトラクタ構造をつくる。
このモデルでは海馬の文脈情報と感覚入力のキーパターン情報が長期記憶の入力となります。前述の実験結果に基づいて、感覚情報入力と海馬情報入力の間の時間関係に依存したタイミング窓を一致性検出回路として導入しています。このモデルの計算機シミュレーション結果によれば、海馬の文脈情報の導入により引き込み領域が拡大するとともに、書き込みおよび読み出し時間が著しく改善され、記憶パターンの正解率も向上しました。また、引き込み領域の拡大は、条件刺激音を聴かせた場合のみ聴覚野の応答面積の有意な増加がみられた実験結果を説明していると考えます。
1皮質−海馬系の情報の相互作用(実験)
大脳皮質における感覚皮質と海馬の情報の相互作用 (Ide et al.2006)
記憶の書き込みは次の2つの過程から成っていると考えられます。第1の過程は出来事などの情報を時空間の文脈として蓄える短期記憶の過程であり、海馬にその機能が存在すると考えられています。 第2の過程は海馬での一時的な短期記憶を必要に応じて安定して蓄える長期記憶の過程であり、皮質(連合野)存在すると考えられています。そこには文脈の類似性や関連性に基づいた記憶構造が存在しています。従って、海馬‐皮質系の記憶書き込み過程では2段階の学習過程が存在し、海馬と皮質ではその機能に適した学習則が存在しています。 しかしながら、海馬‐皮質間の情報の動的な相互作用に関する記憶や学習のメカニズムについては、あまり良く分かっていないのが現状です。
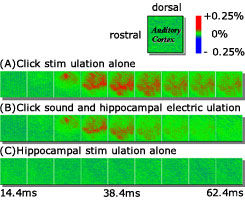
図1 海馬刺激による聴覚野応答の抑制
本研究では海馬CA1と大脳皮質聴覚野の相互作用を調べるため、麻酔下のモルモットの海馬CA1に電気刺激、聴覚に音を提示して大脳皮質聴覚野に生じる応答を膜電位感受性色素(RH795)による光計測法を用いて記録しました。 海馬CA1へは双極のタングステン電極を刺入し、0.19 mAで100・sのパルスによって電流刺激を与えました。その結果、海馬CA1への電気刺激と音刺激のタイミングに依存して聴覚野に生じる神経活動が修飾されることが明らかとなり、その応答は通常(音刺激単独の場合)に比べて抑制されることが分かりました(図1)。この現象は、聴覚皮質に感覚情報と海馬情報の間にタイミングに依存したゲーティング(タイミング窓)が存在することを示しています。
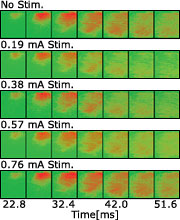
図2 海馬刺激強度の違いによる聴覚野応答への効果
さらに、海馬CA1活動の強さの違いによって聴覚野の応答に異なった影響が生じるかを調べるため、海馬CA1の電流刺激強度を0.19mAから0.76mAまで変化させて聴覚野の応答を比較しました。 電流強度を変化させることは海馬CA1内の活動するニューロンの数を変化させることに相当します。この結果、電流強度が0.38mA以下では海馬への刺激は聴覚野の応答に対し抑制性に働きますが、0.57mA以上では興奮性に働くことが分かりました(図2)。このように海馬CA1の活動規模によって聴覚野の応答に対し抑制性と興奮性の修飾が生じることは、海馬から大脳皮質聴覚野への情報を書き込む際に、より能力の高い情報処理が可能であることを意味しています。 また、この応答変化を詳しく解析した結果、海馬刺激による聴覚野応答の修飾は空間的に一様ではなく、聴覚野の腹側より背側で応答変化がより大きいことが分かりました。
2海馬‐皮質モデルの記憶の理論モデル(理論)
海馬と皮質記憶システムの間の相互作用 (Pan and Tsukada, 2006)
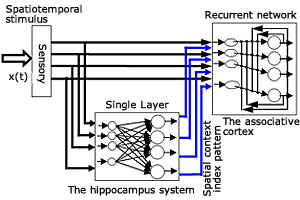
図3 海馬-皮質記憶系の構造
一般に、海馬‐皮質系回路網は、記憶の書き込みと読み出しのための特別な構造をしていることが知られています。海馬は、短期記憶の保持の働きを担っていて、一方大脳皮質は長期記憶の保持の働きを担っています。いかに海馬の短期記憶が皮質の長期記憶に変換されるのでしょうか。我々は、生理学的証拠に基づき海馬‐皮質系記憶システムの理論モデルを提案しました(図3)。このモデルは、感覚システム、海馬(短期記憶)、連合皮質(長期増強)の3つのコンポーネントからなります。1シリーズのキーコード(ローカルな情報)が感覚システムから連合皮質に与えられ、一方海馬それらの文脈情報(グローバルな情報)が連合皮質に与えられます。この2入力が連合皮質で動的に相互作用を行います。そしてこのとき連合皮質のニューロンは時間的一致検出器として働きます。皮質の神経回路網は記憶情報を一致時間窓(the coincidence window)を通して記憶情報を獲得し、最後にアトラクタの形でそれを記憶します。このローカル−グローバル情報によって、連合皮質の記憶の蓄積場所がデザインされ、記憶の書き込みと読み出しの情報処理が実現可能となります (図4 and 図5)。
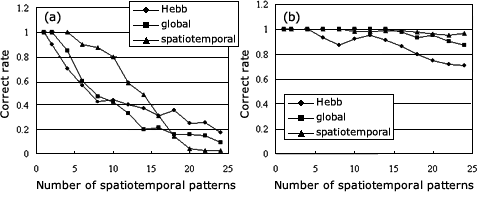
図4 The capability of storing patterns in the associative cortex. (a) The memory system without hippocampus, and (b) the system with hippocampus. The three curves represent different learning rules: the Hebbian learning rule (the circle-curve), the global Hebbian learning rule (the square-curve) and the spatiotemporal learning rule (the triangle-line).
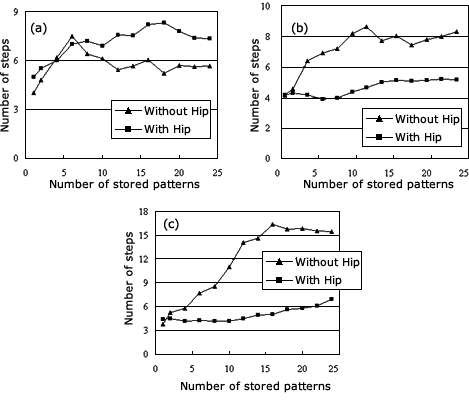
図5 The courses of retrieving patterns from the cortex under the three learning rules. (a) The Hebbian learning rule, (b) the global Hebbian learning rule and (c) the spatiotemporal learning rule. In these figures, the triangle-curve is the result from the memory system without the hippocampus, while the square-curve is the data from the memory system with the hippocampus.
- 【主要業績】1.時空間学習則とその実験的検証
- 【主要業績】2.感覚野(皮質)−海馬の相互作用と記憶の理論モデル
- 研究業績リスト
- グループ研究者紹介
- 【フォーラム】玉川ダイナミックブレインフォーラム
- 【フォーラム】COE若手の会