神経計算論・工学応用研究グループ > 主要業績2
 2.シナプス可塑性が神経細胞の選択性や局所回路の機能に及ぼす影響
2.シナプス可塑性が神経細胞の選択性や局所回路の機能に及ぼす影響
〜シナプスレベルの現象と局所回路レベルの現象をつなぐ〜
STDP[主要業績1]に基づいて、スパイクの相対的タイミングのみで記述できる単純なシナプス可塑性ルールが導かれます。このSTDPルールの計算論的な意義が世界中で検討されてきました (Song et al. 2000;Song and Abbott 2001; van Rossum et al. 2000; Rubin et al. 2001; Gqutig et al. 2003; Toyoizumi et al. 2005)。我々もこの問題に取り組み、Fokker-Plank方程式を用いて、STDPルールにより獲得されるニューロンの選択性の性質を調べる理論的枠組を提供し(Cateau and Fukai 2003)、様々なタイプのSTDPルールがニューロンの選択性に及ぼす影響を調べました(Sakai et al. 2004)。また、局所回路のモデルにSTDPルールを適用し、組織化される局所回路の性質を調べました (Kitano et al. 2002,2002,2003,2004; Kang et al. 2004; Sakai 2005)。
我々が構築した理論的枠組は以下の想定を必要としています。
- 多数のニューロンから入力を受けている1つのニューロンを考える。
- 送信側の各ニューロンは、変動する発火率にしたがってランダムにスパイクを送っています。
- 送信側の変動発火率パターンは同時刻の相関があるが、時間遅れのある相関はない。
- シナプス強度変化は十分にゆっくりとしており、スパイクとスパイクの間の間隔程度の間には、シナプス強度一定と仮定してもよい。
これらの想定のもとで、STDP学習則で十分学習を行った後の定常状態で、その受信側のニューロンが受けているシナプスの強度分布を数値的に計算することができます。もし、さらに以下の仮定をおくと、統計的な平均操作が一部ラプラス変換となるため、シナプス強度分布の解析解を得ることができます。
- 送信側のニューロンの発火率は一定であります。
- 受信側のニューロンは、Leaky integrate-and-fire モデルに従う。
- STDPのタイミング依存性が、シナプス前-後の順、シナプス後-前の順、それぞれ、指数関数で近似できる。
この枠組みを用いて、STDPの初期強度依存性がシナプス強度の定常分布に重大な影響を及ぼすことがわかりました。シナプス増強と減弱では、非対称な初期強度依存性を示すことが報告されています。我々は(Sakai et al. 2004) 様々な形の非対称な初期強度依存性で、どのようなシナプス強度分布を示すか、調べました(図1)。
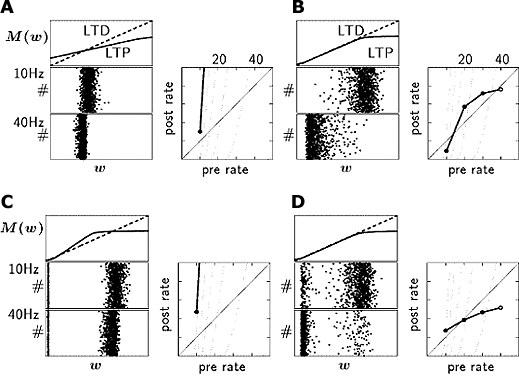
図1 様々なパターンの初期強度依存性を持つ場合のSTDP学習則によって獲得されるシナプス強度分布のパターン。1つのニューロンがそれぞれ一定の発火率でランダムに発火する1000個のニューロンからシナプス結合を受けている。各グラフ(A-D)の関数 M(w) は、シナプス増強(実線)とシナプス減弱(破線)に用いた初期強度依存性を示している。十分に学習し、シナプス強度分布が定常状態になった後のシナプス強度分布は、ある初期強度依存性 M(w) の組み合わせのときには単峰性の分布を示し(A,B)、また別の初期強度依存性 M(w) の組み合わせのときには双峰性の分布を示している(C,D)。双峰性の分布はシナプス間に競合が働いていることを示している。シナプス増強・減弱の初期強度依存性がある区間でバランスが取れている場合(B,D)、STDP学習則は、入力側の発火率の変化に対して受信側の発火率を調節する機能をもつ。入力側の発火率を10Hzから40Hzに上げると、シナプス強度分布が小さいほうに偏り、受信側の発火率の変化を和らげている。一方、シナプス増強・減弱の初期強度依存性がほとんどの区間でバランスが取れていない場合(A, C)、入力側の発火率が少し変化しただけで、受信側の発火率が発散してしまう。
我々は、また、空間的に配置したモデルニューロン集団が局所的に互いに結合しており、入力ニューロン集団から順方向性の結合を受けているような回路にSTDP学習則を適用しました。このような回路は、感覚系のトポロジカルマップを再現するためによく用いられる構造です。このSTDP学習則に基づいたトポロジカルマップモデルは、トポロジカルマップによる情報表現を獲得できることが既に報告されています(Song and Abbott 2000)。我々のグループでは(Sakai 2005)、このSTDPトポロジカルマップモデルが、ある種の多重表現を獲得し、側頭葉IT野における情報表現の性質をよく再現することを示しました。側頭葉IT野では、光計測などで、皮質上の活動領域を観測すると、物体の回転に応じて活動領域が連続的に移動することが知られており、トポロジカルマップの特性を持っていることがわかります。しかし、電極で1つ1つのニューロンの選択性を調べると、最適に応答する図形とは全く異なる図形にも強く応答し、またごく近傍に存在するニューロンでも選択性のパターンが全く異なっていることがよく観測されます。一見矛盾するような、この側頭葉IT野の情報表現がどのように整合し、どのようなメカニズムで獲得されているのか、これまで謎とされてきました。我々が示したSTDPトポロジカルマップモデルでは、入力パターン集合の中の連続的な特徴量が、活動するニューロン集団の中心位置で表現され、入力パターン集合の中の離散的な特徴量が、その活動するニューロン集団の中の詳細な活動パターンで表現されるという多重表現を獲得しています(図2)。したがって活動領域を調べれば、トポロジカルマップの特性を示していますが、全く異なる入力にも反応し、その違いが詳細な活動パターンで表現されているため、隣のニューロンでも全く異なる選択性を示すことがあります。側頭葉IT野の表現と整合しています。自然界で動物がよく目にする物体の画像は、あらゆる画像の中で極めて限定されたものであり、画像空間の中でクラスター状に分布しているはずであり、このような入力構造を反映して、ある種の多重トポロジカルマップが獲得されていると解釈できます。
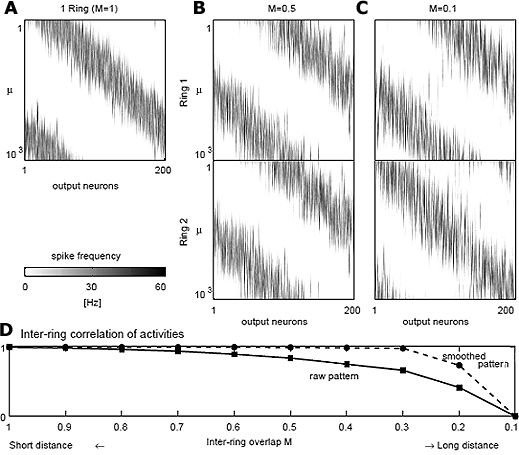
図2 STDP学習則にもとづいたトポロジカルマップモデルでの多重表現。
A-C: 各入力パターン{ξ1,ξ2, ・・・,η1,η2, ・・・} をランダムな順で呈示し、十分学習した後(10,000秒)、各入力パターンξi (Ring 1, E) と入力パターンηi (Ring 2, E)に対する出力ニューロンのスパイク頻度パターンを測り、グレースケールで表示したもの。リング上の位置に関して、高頻度ニューロン群の中心が連続的に対応していることがわかる。
D: Ring1,2の対応する入力パターンに対する、スパイク頻度パターンの方向余弦の平均(実線)と、近傍40ニューロンでスムージングした平滑スパイク頻度パターンの方向余弦の平均(破線)。入力パターンのリング間距離が離れるにつれ、平滑スパイク頻度パターンには違いがないが、詳細なスパイク頻度パターンに違いが生じている。高頻度ニューロン群の中心位置でリング上の位置を、詳細なスパイク頻度パターンでリングの違いを表現している。
参考文献
- Bi G, Poo M (1998) J. Neuroscience, 18:10464-10472.
- Feldman DE.(2000). Neuron, 27:45-56.
- Froemke RC and Dan Y (2002). Nature, 416:433-438.
- Gutig R, Aharonov R, Rotter S, Sompolinsky H (2003). J. Neurosci. 23:3697-3714.
- Rubin J, Lee DD, Sompolinsky H (2001). Physical Review Letters, 86(2):364-367.
- Song S, Abbott LF (2001). Neuron, 32:339-350.
- Song S, Miller KD, Abbott LF (2000). Nature Neurosci., 3:919-926.
- Toyoizumi T, Pfister J, Aihara K, Gerstner W (2005). Proc. Natl. Acad. Sci. USA, 102:5239-5244.
- 【主要業績】1.シナプス可塑性のメカニズム
- 【主要業績】2.シナプス可塑性が神経細胞の選択性や局所回路の機能に及ぼす影響
- 【主要業績】3.神経細胞の発火特性が局所回路の挙動に及ぼす影響
- 【主要業績】4.マッチング行動のメカニズム
- 研究業績リスト
- グループ研究者紹介