
 �X�V���@2026�N6��19��
�X�V���@2026�N6��19��
- �p����
- �i��ꗗ
- �ԏҊ��̋N��
- ������E���������m�n�i�V���E�u�i��̊��j -�]�ˌn�ԏҊ��̐�������`�w�I�ɗ��Â��錴����-
- �����ꗗ
- �N��ʁ@�{�w�ɂ�����͔|��̃n�i�V���E�u�A�쐶��̃m�n�i�V���E�u�̌����Ɛшꗗ
- �ʐ^�B�e���@
- �A���ւ��̃|�C���g
- �y�̎�ށF�L�@���엿�┭�y���i�܂͕s�v�ł�
- 2�`3���̊Ǘ����
- 4�`5���͔̍|�̃|�C���g
- 5�����{�͔̍|���@
- �ď�̍��
- �H�͔̍|�̃|�C���g
- ���̐���f�f�|���F�ɂȂ����t�͂����Ɏ�菜���܂�
- 10�����{�̊��̃`�F�b�N�i���̒������ɒ��Ӂj
- 11�����E���{�̏�ԁi���N�ւ̊��̈ێ��E�ۑ��Ɍ����āj
- �]�ˌn�A���n�A�ɐ��n�́u�Éԁv�̕ۑ��@















�ԏҊ��̋N���i2026�N5��12������j
1�D�͂��߂�
�ԏҊ��iIris ensata�j�́A���{�̓`�����|�A���̒��ł����ɕ����I�E���j�I���l�������A�]�ˎ��ォ�猻��Ɏ���܂ő����̐l�X�ɐe���܂�Ă��܂����B�e�n�œƎ��Ɉ琬���ꂽ�i��Q�́A�n��̕��y����ӎ��f���A���{�̉��|�����̑��l�����ے����Ă��܂��B
�ԏҊ��ł́A�琬�n�̈Ⴂ�ɉ����A���̓y�n�Ŏp����Ă����ӏܗl�����ԗe�E�ԐF�E�`�ԓI�����ɒ��ڔ��f�����Ƃ����Ǝ��̕����I�w�i������܂��B
- �]�ˁF�Q��č炭�p�����ڂɊӏ܂��镶�� → ���炫�E���L����̉ԗe
- �ɐ��F������Ԃ̐L�W���y���ޕ��� → ����炫�̉ԗe
- ���F��ւ𐳖ʂ���[���Ɋӏ܂��镶�� → ��ւŐ������ԗe
- ����F�쐶�I�`�����c�����f�p�ȉԗe
���̂悤�ɁA�n�敶���Ɗӏܗl�����i��`�����̂��̂�����Â��Ă����_�ɁA�ԏҊ��̑傫�ȓ���������܂��B �{�z�[���y�[�W�Ŏ����ԏҊ��l�n���i�]�ˁE�ɐ��E���E����j�̐����j����і쐶�m�n�i�V���E�u�̈�`�\���Ɋւ���m���́A2006 �N�̏���ȍ~�A�{�w���p�����Ď��{���Ă����`�Ԋw�E�����w�E���Ԋw�E�����j�����ƁA���q�����w�I��́icpDNA�ESSR�EMIG-seq�EEST�j���������������Ɋ�Â����̂ł���A�����̌����͍����O�ɐ�삯�đ̌n�I�Ɏ��{���ꂽ��s�����ł��B
 �쐶�m�n�i�V���E�u�����̐i�W�ƁA�ԏҊ��N�������̊��
�쐶�m�n�i�V���E�u�����̐i�W�ƁA�ԏҊ��N�������̊��
�ԏҊ��̋N�����Ȋw�I�ɉ𖾂��邽�߂ɂ́A�i��琬�̊�ƂȂ����쐶�m�n�i�V���E�u�̈�`�\���𗝉����邱�Ƃ��s���ł��B
�{�����́A1930�N��̎O�D�w�ɂ��`�Ԋw�I�����������Ƃ��A2006�N�́u�m�n�i�V���E�u�̕ψِ��Ɋւ��錤���i��1��j�v���N�_�Ƃ����A�`�Ԋw�E�����w�E�����j�E���q�����w�icpDNA�ESSR�EMIG-seq�EEST�j ���������������Ƃ��Ĕ��W���Ă��܂����B
 2009�F���сi�C�m�_���j�ɂ�� cpDNA ��́\�{�B�Ƃ͈قȂ� “�嗤�n��” �̑��݂����߂Ď���
2009�F���сi�C�m�_���j�ɂ�� cpDNA ��́\�{�B�Ƃ͈قȂ� “�嗤�n��” �̑��݂����߂Ď���
2009�N�ɂ́A���сi�C�m�_���A�c���Ƃ̋��������j�ɂ��A���{�e�n����ъ؍��Y���܂ޖ쐶�m�n�i�V���E�u�W�c�̒n���I��`�\�����A�t�Α� DNA�icpDNA�j��p���ď��߂ĕ��q���x���Ő�������܂����B
�����A��B�̐^���Ȗ쐶�W�c�̍ޗ������荢��ł��������߁A�؍��Y�̂��u�嗤�n���̔�r�Ώہv�Ƃ��ĉ�͂��܂����B ���̒i�K�ł́A��B�Y�Ɗ؍��Y������ł��邱�Ƃڎ������킯�ł͂���܂��A�؍��Y���{�B�̏W�c�Ƃ͖��m�ɈقȂ��嗤�n���i�쐶�m�n�i�V���E�u�̈�`�I�n���j�ɑ����邱�Ƃ�������܂����B
���̒m���́A��� Mimura et al.�i2024�j�ɂ��S���K�͂̉�͂ƒ��ڂȂ���d�v�Ȋ�ՂƂȂ�܂����B
2010-2011�F�X�E���E�c���E���c�ɂ��SSR��́\ �u�쐶�� → �]�ˌn�v�̘A���������߂Ď���
2010�`2011�N�ɂ́A�X�E���E�c���E���c��ɂ�� SSR�i�}�C�N���T�e���C�g�j��͂��s���A �쐶�m�n�i�V���E�u�ƍ]�ˌn�ԏҊ��̈�`�I�A���������߂ĕ��q���x���Ŏ�����܂����B
���̌����ɂ��A
- �]�ˌn�ԏҊ��͖쐶�m�n�i�V���E�u���琬��Ƃ��Đ�������
- ���ɖ{�B�����̖쐶�W�c���]�ˌn�̈琬��̌��ł���
�Ƃ������N�̓`�����A���߂ĉȊw�I�ɗ��t�����܂����B
���̐��ʂ́A��� Mimura et al.�i2024�j����� Kobayashi and Tabuchi�i2025�j�ւƂȂ���d�v�Ȋ�ՂƂȂ�܂����B
2024�FMimura et al. �ɂ��S��260�����̖ԗ���́\�嗤�n���̒n���I�A�������m��
2024�N�ɂ́A�S��260�����ȏ�̖쐶�m�n�i�V���E�u��ΏۂƂ����ԗ��I DNA ��́iMimura et al., Conservation Genetics�j���s���AMIG-seq �ɂ�鍂�𑜓x�̃Q�m����͂̌��ʁA���{�̖쐶�m�n�i�V���E�u��3�̎�v�Ȉ�`�I�n���igenetic lineages�j �ɕ�����邱�Ƃ����炩�ɂȂ�܂����B
���ɁA��B�Y�ƃ��V�A�Y������́u�嗤�n���i�쐶�m�n�i�V���E�u�̈�`�I�n���j�v�ɑ����邱�Ƃ����q���x���Ŏ�����A���V�A → ���N���� → ��B�ւƘA�������`�I���C���̑��݂����m�ɂȂ�܂����B
���̒m���́AcpDNA��p�������сi2009�j���؍��Y�m�n�i�V���E�u���u�嗤�n���i����B�Ɠ��n���j�v�Ƃ��ď��߂Ĉʒu�Â������ʂ��AMIG-seq�ɂ��Q�m����͂����łɗ��t�������̂ł��B
����ɁA2010-2011�N��SSR��͂��������u�쐶�� → �]�ˌn�v�̘A�����Ƃ��������A�嗤�n�������N�������o�R���ċ�B�ɓ��B�����Ƃ����n���I�E��`�I�A�������A2024�N�̌����ɂ��ŏI�I�Ɋm�肵�܂����B
2025�F�l�n���i�]�ˁE�ɐ��E���E����j�̋N�������ۓI�Ɋm��
2025�N�ɂ́A���ۉ��|�w��ł̔��\���o�āAActa Horticulturae�ɍ]�ˌn�E�ɐ��n�E���n�E����Î�̋N���Ɋւ���2��̘_�����f�ڂ���A�l�n���̐����j�����ۓI�ɏ��F����܂����B ����ɂ��A�ԏҊ��l�n���̋N���͐��E�ŏ��߂ĉȊw�I�ɑ̌n������܂����B
�����ޗ��̏d�v��
�ߔN�A���n���̕ω���J���ɂ���쐶�m�n�i�V���E�u�̎����n�͋}���Ɍ������A���|�i��Ƃ̌��G�ɂ���`�I�e���������������A�^���Ȗ쐶�W�c�̈ێ�������ɂȂ����܂��B
�ÉԂɂ��Ă��A���ω���Ǘ��̐��̕ω��ɂ��ێ�������A���������̐V�ԁi�n���Ԍ��G�i��j�ɒu��������Ă��܂��B
�{�w�ł́A���ω��ȑO��1980�N��ɍ̎悵40�N�Ԉێ����Ă����쐶�m�n�i�V���E�u�ƁA�^�������m�F�����ÉԂ�30�`40�N�ԓ�������ō͔|���Ă����W���̂�p���Č�����i�߂Ă��܂����B �����̍ޗ��͌��݂ł͓��������ōĂё����邱�Ƃ�����A�����̈�`�\���f���������͌��݂ł͍Č����ɂ߂č���ł��B
�{�����̈Ӌ`
�ԏҊ��l�n���̋N�����Ȋw�I�ɐ����ł����̂́A �����ɂ킽��ێ��E�ۑ�����Ă����ÉԂƁA���ω��ȑO�̖쐶�m�n�i�V���E�u�Ƃ����A�ɂ߂ċM�d�Ȍ����ޗ��������Ă������߂ł��B
�{�����́A�O�D�w�ȗ��A��ꐢ�I�Ԃ�ɉԏҊ��̋N���𑍍��I�ɍč\�z���������ł���A����̕ۑ��E���E�����j�����̊�ՂƂȂ���̂ł��B
���p�E�]�ڂɂ���
�{�y�[�W�̐}�\����ѕ��͂̓]�ځE���p�ɂ́A�{�w�L��ۂ̎��O�����K�v�ł��B �܂��A�����p�Ƃ��Ē����ێ����Ă����W�����́A�Č����m�ۂ̊ϓ_������n�E�������s���Ă���܂���B
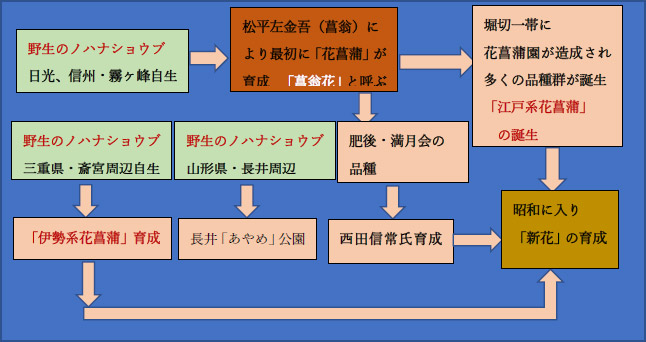
2�D�]�ˌn�ԏҊ��̋N��
���݂̉ԏҊ����Ɍ�����悤�ȍ͔|�i��́u�ԏҊ��v��̌n�I�Ɉ琬�����̂́A�]�ˎ���̏���������i���F�҉��j�ł��B�҉��́A�e�n������肵���쐶�m�n�i�V���E�u�̎�q���]�˂Ŕd�킵�A�I���E�琬���s���܂����B�����̕i��Q�͌�ɑ��̂��āu�]�ˌn�ԏҊ��v�ƌĂ��悤�ɂȂ�܂����B
�]�ˌn�ԏҊ��̋N�����Ȋw�I�ɖ��炩�ɂ��邽�߂ɂ́A �u���������Ⴊ�ǂ̒n��̖쐶�m�n�i�V���E�u���̂Ƃ��ĕi����琬�����̂��v ����肷��K�v������܂��B
�����ɕK�v�ȍޗ��͈ȉ���2�ł��B
- �S���e�n�Ɏ�������쐶�m�n�i�V���E�u
- ���������Ⴊ�琬�����Ƃ����u�҉��ԁv�i�Éԁj
�i1�j2009�F���сi�C�m�_���j�{�c��-cpDNA �ɂ���b���
2009 �N�̏��сi�C�m�_���j�ł́A�c�����̎�E�ێ����Ă����쐶�m�n�i�V���E�u��p�����t�Α� DNA�icpDNA�j������s���A �k�C���E���k�E�{�B�E�O�d�E�؍��̖쐶�W�c�̒n���I��`�\�������߂Đ�������܂����B
���̌����ɂ��A �]�ˌn�ԏҊ��̕�̌�₪�{�B�����ɂ��邱�Ƃ���������܂����B
�i2�j2024�FMimura et al. -MIG-seq�ɂ��S���ԗ����
�S��260�����ȏ�̖쐶�m�n�i�V���E�u��ΏۂƂ���MIG-seq��͂ɂ��A�]�ˌn�ԏҊ��̕�̂��{�B�����i������E�����j�ɂ��邱�Ƃ�������������܂����B
�i3�j2025�FKobayashi and Tabuchi -EST�i�A�C�\�U�C���j�ɂ��ŏI�m��
�Éԁi�҉��ԁj�ƑS���̖쐶�m�n�i�V���E�u�W�c���r���� EST�i�A�C�\�U�C���j����ɂ��A�]�ˌn�ԏҊ��͖�����E�����̖쐶�W�c���琬��Ƃ��Đ����������Ƃ��AEST�i�A�C�\�U�C���j��͂ɂ�蕪�q���x���ōŏI�I�Ɋm�肵�܂����B

Mimura et al.�i2024�j�ƁAKobayashi and Tabuchi�i2025�j�ɂ���Ċm��
3�D�ɐ��n�ԏҊ��̋N��
�ɐ��n�ԏҊ��́A�]�ˎ���̒����`����ɂ����āA�I�B�ˎm�E�g���ܘY�ɂ���Č��݂̎O�d��������ӂň琬���ꂽ�Ɠ`�����Ă��܂��B�������A���̋N���ɂ��Ă͒����c�_�������Ă��܂����B
�i1�j���j�I�w�i�Ɛ�s����
�y��i1967�j�́A�Q�Ό���ɐ��Q���ʂ��č]�ˌn�ԏҊ����ɐ��Ɏ������܂ꂽ�ƍl���A�ɐ��n�͍]�ˌn����h�������Ǝ咣���܂����B ����A�����i1981�j�͎��g�̌�z�����̊ώ@����A�ɐ��n�͍]�ˌn����n�Ƃ͈قȂ��`�I�w�i�����\�����L�^���܂����i�c��, 2016�j�B
�i2�j�{�w�ɂ�鑍�������i�`�ԁE�����E���ԁE���q�j
�{�w�ł́A
- �`�Ԋw�I�����i�ԗe�E�O�Ԕ�Ђ̊p�x�E�Ԍa�j
- �����w�I�����i�J�Ԏ��̐L�W�p�^�[���j
- ���Ԋw�I�����i�����n�̊������j
- ���q�����w�I��́icpDNA�EEST �A�C�\�U�C���j
�𑍍��I�Ɍ������A �ɐ��n�ԏҊ��̕�͎̂O�d���E�{�Ɏ�������m�n�i�V���E�u�ł������Ƃ𖾂炩�ɂ��܂����i���сE�c��, 2020�GTabuchi and Kobayashi, 2025�j�B
�i3�j�����ɗp�����ޗ��i�i��E�쐶�W�c�j
�����ɗp�����ޗ��́A �ɐ��n�ԏҊ��͔̍|�i��ƁA�쐶�m�n�i�V���E�u�i���݂͈�`�q�N�H�ɂ�茤��������j���܂ށA�ȉ���3�Q�ɕ��ނ���܂��B
- �@�ɐ��Éԁi40�i��j
�ɐ��E����n���ő�X�ۑ�����Ă����ÉԁB�]�ˁ`�������̌`����ێ������^���N���[���ł���A�ɐ��n�̈�`�I�w�i��������B��̍ޗ��ƂȂ�܂��B
- �A�ɐ��n�u�V�ԁv�i9�i��j
���a���ɕy�쎁���琬�����i��Q�B �ɐ��ԗe�������A�ÉԂƂ͋�ʂ����B��`�I�w�i���s���m�ł��邽�߁A�{�����̕��q��͂ɂ͗p���Ă��܂����B
- �B��r�ΏƂƂ��Ă̍]�ˌn�ԏҊ��i45�i��j
�O���`�Ԃ��ɐ��n�Ɨގ�����]�ˌn�i���I������r�Ɏg�p�B
�i4�j�쐶�m�n�i�V���E�u�i�{�E�ɉ�E���m�j
�쐶�m�n�i�V���E�u�Ƃ��ẮA
- �O�d���E�{�̎�����
- �ɉ�n���̎�����
- ���m���̎�����
��p���܂����B �����̑����͍��w��V�R�L�O���ł��邽�߁A �Ǘ����ǂ̋��č̎�E�������s���܂����B�����ɗp�����̐��́A �����c���܂߂Ė� 1000 ���ɋy�т܂��B
�i5�j�������ʂƗ��j�I�m���Ƃ̐�����
�`�ԁE�����E���ԁEcpDNA�EEST �̂��ׂĂ���v���A �ɐ��n�ԏҊ��̕�͍̂{�̖쐶�m�n�i�V���E�u�ł������Ƃ������x������܂����B
4�D���n�ԏҊ��̋N��
���n�ԏҊ��́A�]�ˎ���̓V�۔N�ԂɁA���ˎ�E�א�Č삪����������i�҉��j�ɉԏҊ��̕i����n�����肵�����ƂɎn�܂�܂��B �����̋L�^�ɂ́u��O�s�o�������ɏ҉��Ԃ�������v�Ɩ��L����Ă���A���n�ԏҊ��̕i��Q�́A�쐶�m�n�i�V���E�u�ł͂Ȃ��A�]�ˌn�ԏҊ��̒��ł����ɏ҉��ԁi����������琬�i��j���̂Ƃ��Đ����������Ƃ����j�I�ɗ��t�����Ă��܂��B
�{�w�ł́A���̗��j�I�L�^�q�����w�I�Ɍ����邽�߁A �҉��ԂƔ��n�i��̈�`�I�W�� EST�i�A�C�\�U�C���j��͂ɂ�薾�炩�ɂ��܂��� �i�c����, 2023�GKobayashi and Tabuchi, 2025�j�B
�i1�j�����ɗp�����ޗ�
�{�����Ŏg�p�����ޗ��́A���ׂč͔|�i��ł���A�ȉ���3�Q�ɕ��ނ���܂��B
- �@�]�ˌn�i��i�҉��ԁj25�i��
����������i�҉��j���琬�����i��Q�B ���n�̕�̂Ƃ��ė��j�I�ɋL�^����Ă���B
- �A���n�u������i��v
���E�F�{�鉺�ŁA�Ԃ̑g�D�u������v�ɂ���č͔|�E�ێ����ꂽ�i��Q�B �]�ˌn�i�҉��ԁj����ɉ��ǂ��ꂽ�Ɠ`������B
- �B���n�u���c���琬�i��v
��������A�����������ł��������c�M�펁�����l�ň琬�����i��Q�B ������i�����ɓƎ��ɉ��ǂ������́B
�i2�j�������ʁF���n�̐����ߒ������q���x���Ŋm��
���q�����w�I��́iEST �A�C�\�U�C���j�ɂ��A�ȉ��̂��Ƃ����炩�ɂȂ�܂����B
- �@���E������i���82�����҉��ԗR��
���n�̎�v�i��́A �҉��Ԃ��̂Ƃ��Đ����������Ƃ����q���x���ŗ��t�����܂����B
- �A���Ɂu�F���v�u�����N�v�ȂǓ���̏҉��Ԃ������֗^�i78.4���j
���n�̉ԗe�`���ɑ傫����^�����҉��Ԃ����肳��܂����B
- �B���c���琬�i���81�� ��������i��R��
���c�M�펁�́A ��ォ�牡�l�Ɉڂ�Z���������i�����Ɉ��𑱂������Ƃ����q���x���Ŋm�F����܂����B
�i3�j���n�̕��ށi�{�w�̐����j
���A���n�ԏҊ��͂��̍��ȉԗe����S���I�Ȑl�C���W�߂܂����i�c��, 2016�j�B �������A���j�I�L�^�ƕ��q��́i�҉��ԁE������i��E���c���琬�i��̔�r�j�Ɋ�Â��A�{�w�ł͈ȉ��̂悤�ɕ��ނ��Ă��܂��B
- ����
→������i��A����ѐ��c�M�펁�琬�i��B
���j�I�ɘA�������n���ł���A���n�̊���\������B
- �V�ԁi���ԗe�j
→���a���Ɍ��c�`�j���E�����G�ꎁ���琬�����i��Q�B
���ɓƗ����Ĉ琬���ꂽ�i��Q�ł���A�ÉԂƂ͌n���I�w�i���قȂ�B
���̕��ނɂ��A���n�̗��j�I�A�����ƈ�`�I�w�i�m�ɋ�ʂ��邱�Ƃ��\�ƂȂ�܂����B
5�D�R�`���E����n���̉ԏҊ��Q�̋N��
�R�`���E����n���ɂ́A�u����v�ƌĂ��Ǝ��̉ԏҊ��i��Q�����݂��܂��B �{�w�ł́A������ �u����i��O�j�v �ƕ\�L���Ă��܂��B
�����̕i��Q�͍͔|��ł���Ȃ���A
- �쐶�m�n�i�V���E�u�ɋɂ߂ċ߂��Ԍ`�E�ԐF
- �����̎�q���������鐶�ԓI����
- �J�Ԋ����쐶�Ɠ���
�Ƃ����������������A �쐶�m�n�i�V���E�u�ɔ��ɋ߂�������ێ����Ă���_���傫�ȓ����ł��B
����s�u����ߌ����v�ɕۑ�����Ă��邱���̕i��Q�́A ��������ɋ��c���������A���ѐՂɋߗׂ́u����߁v��A���t�������Ƃ��R���Ƃ���Ă��܂��B
�i1�j����i��Q�̋N���Ɋւ��鏔��
����i��Q�̋N���ɂ��ẮA����܂ł�
- �]�ˌn�ԏҊ����������܂ꂽ�Ƃ����
- �]�ˌn�i�킪�n��̖쐶�m�n�i�V���E�u�ƌ��G���Đ��������Ƃ���� �ȂǁA�����̐����������Ă��܂����B
�����͂�������A����n���̉ԏҊ����쐶�I�`���������ێ����Ă��邱�Ƃ��琶�܂ꂽ�����ɉ߂����A�Ȋw�I�Ȍ��͂���܂ōs���Ă��܂���ł����B
�i2�j2009�F���тɂ�� cpDNA ���
2009�N�A���сi�c���Ƃ̋��������j�ɂ��A����s���ӂ̖쐶�m�n�i�V���E�u�� cpDNA �^���A���n��Ƃ͈قȂ�Ǝ��̈�`�^�������Ƃ�������܂����B
���̉�͂ɂ��A ����n���̖쐶�W�c���Ɨ�������`�I�w�i�������Ƃ����߂Ė��炩�ɂȂ�܂����B���̒m���́A��̕��q��͂̊�ՂƂȂ�܂����B
�i3�j�{�w�ɂ�镪�q�����w�I��́iEST �A�C�\�U�C���j
�{�w�ł́A
- ����s���ӂɎ�������쐶�m�n�i�V���E�u
- ����u����ߌ����v�ɕۑ�����Ă���i��Q
��ΏۂɁAEST�i�A�C�\�U�C���j��͂��s���܂����B
�����ޗ��͈ȉ���2�Q�ł��B
- �@����s���ӂɎ�������쐶�m�n�i�V���E�u
���ω��ȑO����ێ�����Ă����̂𒆐S�ɉ�́B
- �A����u����ߌ����v�ɕۑ�����Ă���i��Q
�i�햼���t�^����Ă���͔|�́B
�i4�j�������ʁF����i��Q�́u�n���̖쐶�W�c�R���v�ł���
��͂̌��ʁA ����i��Q�́A����s���ӂɎ�������쐶�m�n�i�V���E�u���m�����R���G���A ���̒�����I������ĕi�햼���t�^���ꂽ���̂ł������Ƃ����炩�ɂȂ�܂����B
�܂�A
- �]�ˌn�ԏҊ�����������
- �ɒB�Ƃ��Q�Ό��Ŏ����A����
- �]�ˌn�Ɩ쐶�����G����
�Ƃ��������͎x�����ꂸ�A ����i��Q�́u�n���̖쐶�W�c���̂Ƃ���Ɨ��n���v�ł������Ƃ��Ȋw�I�Ɋm�肵�܂����B�Ȃ��A�ꕔ�̕i��ł́A�㐢�͔̍|�j�̒��ō]�ˌn�i��̈�`�q�����������\������������܂����A����i��Q�S�̂̐����j�����E������̂ł͂���܂���B
�i5�j2026�N8���F���ۉ��|�w��Ŕ��\�\��
�{�������ʂ́A 2026�N8���ɊJ�Â���鍑�ۉ��|�w��iInternational Horticultural Congress�j�ɂĔ��\�\���ł��B
����ɂ��A
- 2009�F����쐶�W�c�̓Ǝ����icpDNA�j
- 2026�F����i��Q�̋N���iEST�j
�����ۓI�ɕ���A ����i��O�j�̋N���������ɍ��ۊw��Ŋm����������ƂɂȂ�܂��B
6�D�ԏҊ��̕i�퐫�����̂܂Ƃ�
�{�w�̌������ʂ���A �]�ˌn�ԏҊ��A�ɐ��n�ԏҊ��A���n�ԏҊ��A����ђ���i��Q�i��O�j�̗R���́A �ȉ��̂悤�ɑ̌n�I�ɐ������邱�Ƃ��ł��܂��B
- �]�ˌn�ԏҊ��F������E�����̖쐶�m�n�i�V���E�u���̂Ƃ��Đ���
- �ɐ��n�ԏҊ��F�O�d���E�{�̖쐶�m�n�i�V���E�u���̂Ƃ��Đ���
- ���n�ԏҊ��F�]�ˌn�́u�҉��ԁv���̂Ƃ��Ĕ��ʼn���
- ����i��Q�i��O�j�F����s���ӂ̖쐶�m�n�i�V���E�u���̂Ƃ��Ď��R���G�Ő���
�����l�n���̐����j�́A 2009 → 2024 → 2025 → 2026 �Ƒ����{�w�̌����ɂ��A���E�ŏ��߂ĉȊw�I�ɑ̌n������܂����B
����̉ۑ�́A �����͔̍|�i��������ɉi�N�I�Ɉێ��E�ۑ����Ă������A �����Ĉ琬�̊�ՂƂȂ����쐶�m�n�i�V���E�u�̕ۑS���ǂ̂悤�ɐi�߂邩�ɂ���܂��B
�i1�j�����ΏۂƂ����͔|�i��i�Éԁj�̈���
�{�w�ł́A�]�ˎ���Ɉ琬���ꂽ�]�ˌn�E���n�E�ɐ��n�̌ÉԁA ����ђ���n���̕i��Q�ɂ��āA �������ɂ���Čp������Ă����݂̂̂������ΏۂƂ��܂����B
�������i�N���[���j�ɂ���Ĉێ����ꂽ�i��́A �琬�����Ɠ���̌`���i��`�q�j��ێ����Ă������߁A �{�w�ł�30�N�ȏ�ɂ킽��J�Ԃ��m�F���A �Õ����Əƍ��̂����A ���ׂĂ̐A���튯�ɂ����Ĉ�v���銔�݂̂�ۑ����Ă��܂��B
����A ��q�ɐB�ɂ�蓯���ŗ��ʂ��Ă��銔�ɂ��ẮA ��`�I�ψق������Ă���\�������邽�ߌ����Ώۂ��珜�O ���Ă��܂��B
�i2�j�쐶�m�n�i�V���E�u�̕ۑS�ƌ����ޗ��̏d�v��
1986�N�����A�S���ɂ͖� 260 �n��̃m�n�i�V���E�u�����n���m�F����Ă��܂����B �������A�n���K�͂̊��ω���J���̉e���ɂ��A���݂ł͂��̑����������A�����킸�������c���Ă��܂����B�Ȃ��A���݂킸���Ɏc����Ă���m�n�i�V���E�u�����n�ɂ��Ă��A�ߔN�̊��ω���l�דI�ȐA�͂ɂ��A��`�q�N�H��������\�����w�E����Ă��܂��B ���̂��߁A���ω��ȑO�̈�`�\����ێ����錤���ޗ��̉��l�́A����܂��܂����܂�ƍl�����܂��B
�{�w�ł�1986�N���A�NJ����ǂ̋��č̎悵���쐶�����������ɂ��ێ��E�ۑ����A �쐶��̕ۑS�ɂ��p���I�Ɏ��g��ł��܂��B
�����̍ޗ��́A���ω��ȑO�̈�`�\����ێ�����ɂ߂ċM�d�Ȍ��������ł���A ���݂ł͓��������ōĂё����邱�Ƃ͂قڕs�\�ł��B
�i3�j�{�����̈Ӌ`
�{�����́A�O�D�w�i1930�N��j�ȗ��A��ꐢ�I�Ԃ�ɉԏҊ��l�n���̋N���𑍍��I�ɍč\�z�������̂ł���A����̕ۑ��E���E�����j�����̊�ՂƂȂ鐬�ʂł��B
�܂��A2009�icpDNA�j→ 2024�iMIG-seq�j→ 2025�iEST�j→ 2026�i���ۊw��\�j�Ƒ�����A�̌����ɂ��A�ԏҊ��̐����j�����E�ŏ��߂ĉȊw�I�ɑ̌n������邱�ƂɂȂ�܂��B
�i4�j�Љ�I�]���ƎЉ�I����
�{�����́A�쐶�m�n�i�V���E�u�̕ۑS�Ɠ��{�̉��|�����̌p���Ɋւ��d�v�Ȑ��ʂƂ��āA�Љ�I�ɂ������]������Ă��܂��B
2026�N�ɂ́A���{�o�ϐV���iScience ����j�ɂ����āA�c���r�l�u�ۑS�œ��{�炵����� �\�w�Ԃ��݁x���A���Ɋ�@�v�i2026�N2��1���j���f�ڂ���A�쐶�m�n�i�V���E�u�̕ۑS�̏d�v���ƁA�{�������ʂ����������L���Љ�ɏЉ��܂����B �܂��A�{�����̐��ʂ͑��̕@�ւ��厏�ł����グ���A�ԏҊ��̋N�������Ɩ쐶��ۑS�̏d�v�����Љ�I�S���W�߂Ă��܂��B
7�D�Q�l����
A�D�w�p�����i���ǘ_���E���ۉ�c�E�w�p���j
Acta Horticulturae�i���ۉ��|�w��j
- Kobayashi, T. and Tabuchi, T. (2025). Characteristics and appreciation style of Japanese irises (Hana-shobu) 1. Edo-group and Higo-group. Acta Horticulturae, 1434, 379-386.
https://doi.org/10.17660/ActaHortic.2025.1434.52 �i���o�F2024�N4�� ���ۉ��|�w��j - Tabuchi, T. and Kobayashi, T. (2025). Characteristics and appreciation style of Japanese irises (Hana-shobu) 2. Ise-group. Acta Horticulturae, 1434, 367-378.
https://doi.org/10.17660/ActaHortic.2025.1434.51 �i���o�F2024�N4�� ���ۉ��|�w��j
WOLZ / WOTZ Book�i���ۉ��|�w����j
- Kobayashi, T. and Tabuchi, T. (2024). Characteristics and appreciation style of Japanese irises (Hana-shobu) 1. Edo-group and Higo-group. WOLZ Book, 98. International Society for Horticultural Science.
- Tabuchi, T. and Kobayashi, T. (2024). Characteristics and appreciation style of Japanese irises (Hana-shobu) 2. Ise-group. WOTZ Book, 99. International Society for Horticultural Science.
Conservation Genetics�i���ێ��j
- Mimura, M., Takeuchi, M., Kobayashi, T., Hirota, S. K., Suyama, Y., Yamaguchi, H., and Tabuchi, T. (2024). Genetic diversity in the traditional cultivars and wild ancestor (Iris ensata Thunb.) of near-threatened Japanese iris. Conservation Genetics, 25, 1079-1086.
https://doi.org/10.1007/s10592-024-01625-7
���|�w�����i���|�w��j
- �c���r�l�E��������E�����c�m�E���эF���i2023�j�D�G�X�e���[�[�A�C�\�U�C����p�������n�n�i�V���E�u�̋N���Ɋւ��錤���D���|�w����, 22(1), 156�D
- ���эF���E�a�c���E�l�������E�c���r�l�i2017�j�D�A�C�\�U�C����͂��猩���n�i�V���E�u�̋N���\�m�n�i�V���E�u�Ƃ̔�r�\�D���|�w����, 16(1), 412�D
- ���эF���E�l�������E�c���r�l�i2017�j�D�m�n�i�V���E�u�̕ψِ��Ɋւ��錤���i��37��j���쌧��������ӂɎ�������̌Q�̊O���`�ԓI�����D���|�w����, 16(1), 413�D
- �X�h�h�E���t���E�c���r�l�E���c���i2011�j�DSSR��͂ɂ��n�i�V���E�u���|�i��Q�ƃm�n�i�V���E�u�W�c�Ƃ̊W�ɂ��āD���|�w����, 11(1), 209�D
- �X�h�h�E���t���E�c���r�l�E���c���i2010�j�DSSR�}�[�J�[�̊J���Ɖ�́D���|�w����, 9(1), 397�D
- �c���r�l�E������E�ݑ�L�I�E�������E������E���{�a�O�i2010�j�D�m�n�i�V���E�u�̕ψِ��Ɋւ��錤���i��17��j�O�Ԕ�Ђ̌`�Ԃ̒�ʓI�]���D���|�w����, 9(1), 437�D
���ۉ�c�iIHC�j
- Handa, T., Mori, T., and Tabuchi, T. (2010). Development of microsatellite markers in Iris ensata (Iridaceae). 28th International Horticultural Congress, 248.
��w�I�v
- Chino, N., Kobayashi, T., and Tabuchi, T. (2020). Characteristics of the Japanese wild iris mutant showing white color on the rim of the outer perianth found around the Shirakami Sanchi. Shirakami Kenkyu, 14, 49-64.
�w�ʘ_��
- ���яˎq�i2009�j�D�t�Α�DNA�Ɋ�Â��n�i�V���E�u�i��Q�̐����ߒ��̐���D�C�m�_���C�}�g��w��w�@�������Ȋw�����ȁD
B�D��ʎ��E���ЁE�Љ�I�]���i�V���E�T�C�G���X���j
- �c���r�l�i2026�j�D�ۑS�œ��{�炵����� �\�u�Ԃ��݁v���A���Ɋ�@�D���{�o�ϐV���iScience ����j�C2��1���D
- �c���r�l�i2025�j�D�A���̉\������̃R���N�^�[�D�ʍ����o�T�C�G���X�u�A�������߂� �������p�ɔ�߂�ꂽ��v�C70-73�D�i���o�F���o�T�C�G���X 2024�N8�����j
- �c���r�l�i2024�j�D�A���̉\������̃R���N�^�[�D���o�T�C�G���X 2024�N8�����F10-13�D
- �c���r�l�i2019�j�D�ԏҊ��̕i�핪���̗��j�ƁA�i��琬�̊�ɂȂ�������̃m�n�i�V���E�u�D���s���|�C103�C46-50�D
- �c���r�l�i2016�j�D�Ԃ̕i����ǂ̓��{�j�D�w�ԏҊ��x�i�ēc���v�ďC�j�C231-257�D�I���فD
- �c���r�l�i2016�j�D�ԏҊ��̕i�핪���̗��j�ƁA���̖쐶��m�n�i�V���E�u�D���|�t�H, 586�D
- �c���r�l�i2014�j�D�`�����|�A���̃i�V���i���R���N�V�����D�ÓT���|�A���̉ԏҊ��\���̋N���ƂȂ����m�n�i�V���E�u�̕������E��`�����Ƃ��Ẳ��l�D�i�����j���{�A��������D
- �c���r�l�i2009�j�D�ŐV�_�ƋZ�p �ԙ� vol.1�w�m�n�i�V���E�u�x�D�_�����C319-324�D